





Abstract
In the Brassicaceae, the acceptance of compatible pollen and the rejection of self-incompatible pollen by the pistil involves complex molecular communication systems between the pollen grain and the female reproductive structures. Preference towards species related-pollen combined with self-recognition systems, function to select the most desirable pollen; and thus, increase the plant's chances for the maximum number of successful fertilizations and vigorous offspring. The Brassicaceae is an ideal group for studying pollen–pistil interactions as this family includes a diverse group of agriculturally relevant crops as well as several excellent model organisms for studying both compatible and self-incompatible pollinations. This review will describe the cellular systems in the pistil that guide the post-pollination events, from pollen capture on the stigmatic papillae to pollen tube guidance to the ovule, with the final release of the sperm cells to effect fertilization. The interplay of other recognition systems, such as the self-incompatibility response and interspecific interactions, on regulating post-pollination events and selecting for compatible pollen–pistil interactions will also be explored.
Introduction
The acceptance of compatible pollen and the subsequent steps leading to successful fertilization is a complex and co-operative process between the pollen and the receptive pistil. In the crucifer family (Brassicaceae), once a compatible pollen grain lands on the stigma, pollen grain adhesion occurs, followed by foot formation, pollen hydration, and germination. The pollen tube emerges and penetrates the stigmatic surface. It is then guided through the stigma, style, and septum, and finally onto the funiculus to enter the micropyle of the ovule where fertilization can occur (Fig. 1). The sequential events from pollen adhesion to the path of pollen tube growth through the pistil to the ovule for fertilization have been carefully documented at the ultrastructural level in Brassica spp. and Arabidopsis thaliana (Hill and Lord, 1987; Elleman and Dickinson, 1990; Elleman et al., 1992; Kandasamy et al., 1994; Hulskamp et al., 1995,b; Lennon et al., 1998). For all of these steps, there is the basic requirement of proper reproductive tissue formation (reviewed in Blackmore et al., 2007; Colombo et al., 2008; Crawford and Yanofsky, 2008; Punwani and Drews, 2008; Dickinson and Grant-Downton, 2009). In addition to developmental mutants, plants defective in pollen tube growth and guidance have been identified through genetic screens leading to the identification of female and male components regulating post-pollination events (Preuss et al., 1993; Hulskamp et al., 1995,a; Johnson et al., 2004; Boavida et al., 2009). Finally, a number of species in the Brassicaceae are known to have self-incompatibility systems which lead to the recognition and rejection of ‘self’ pollen at a very early stage of pollen–pistil interactions (reviewed in de Nettancourt, 2001; Franklin-Tong, 2008). Thus, there is clearly a complex level of communication taking place between the pollen and pistil to ensure successful fertilization. This review discusses the step-wise mechanisms involved in compatible pollen responses for members of the Brassicaceae, and highlights where species specificity to the pollen–pistil interactions is determined and how the self-incompatibility pathway intercepts the compatible pollen response.
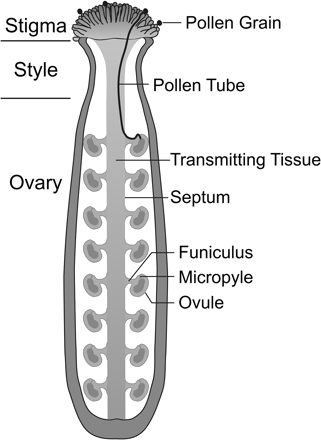
Schematic of a pollinated A. thaliana pistil.
Pollen capture and adhesion to the stigmatic papillae
Members of the Brassicaceae have a dry stigma, which refers to the absence of free-flowing surface secretions; one of the interesting features of this trait is the early selectivity of pollen capture following pollination (Heslop-Harrison and Shivanna, 1977; Dickinson, 1995). Once pollen grains come into contact with stigmatic papillae, only pollen grains recognized as compatible are accepted, thus allowing plants to ignore foreign pollen (Fig. 2). These compatible interactions appear to be confined to species within the family, but clearly can occur beyond the species level (Hulskamp et al., 1995,a). For example, successful pollinations, as measured by pollen tube penetration into the stigma, have been observed in interspecific and intergeneric crosses in the Brassicaceae (Sampson, 1962; Hiscock and Dickinson, 1993; Lelivelt, 1993). However, these interactions are not straightforward as there are other factors governing pollen acceptance within the Brassicaceae. For instance, if present, the self-incompatibility system is activated at the stage of pollen adhesion, and self-pollinations, as well as reciprocal pollinations between plants sharing the same self-incompatibility alleles, are rejected (reviewed in Fujimoto and Nishio, 2007). Furthermore, interspecific or intergeneric pollen acceptance can be regulated by a phenomenon called unilateral incompatibility where pollen from one species is rejected by the pistil of another species, yet the reverse cross is successful (Sampson, 1962; Hiscock and Dickinson, 1993; Takada et al., 2005). For example, no pollen tube penetration was observed on self-incompatible B. oleracea pistils when pollinated with different Brassicaceae species/genera pollen, while self-fertile A. thaliana accepted a wide range of pollen grains following the same survey (Hiscock and Dickinson, 1993). Typically, unilateral incompatibility occurs when the female recipient is self-incompatible and the pollen donor is from a different species that is self-fertile. Consistent with unilateral incompatibility, pollinations between two different self-incompatible species are also typically unsuccessful (Sampson, 1962; Hiscock and Dickinson, 1993). Thus, there appears to be a basic ‘family-wide’ pollen recognition system present in the Brassicaceae, but this recognition can be attenuated in stigmas of self-incompatible plants for interspecific or intergeneric pollen.
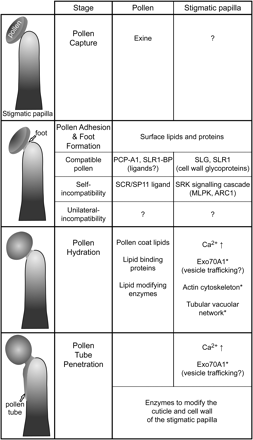
Factors regulating pollen–stigma interactions in the Brassicaceae. Illustrations for the four post-pollination stages are shown, and pollen and stigma factors that are implicated in each stage are listed. Asterisks (*) indicate events that are inhibited or disrupted during the self-incompatibility response. Please see the text for further details and references.
Two studies have specifically examined the early stages of pollen capture and adhesion in interspecific pollinations, and observed that pollen from plants outside the Brassicaceae displayed much lower adhesion to B. oleracea (Luu et al., 1998) and A. thaliana (Zinkl et al., 1999) stigmas as expected. Despite the different approaches used to measure pollen adhesive forces (reviewed in Heizmann et al., 2000), both studies also found that crosses between Brassica spp. and A. thaliana tended to show poor pollen capture and adhesion (Luu et al., 1998; Zinkl et al., 1999). Perhaps, some increased specificity in pollen capture is modulated by different binding affinities at this early stage. However, Luu et al. (1998) surveyed several other Brassicaceae spp. and found high levels of pollen adhesion following interspecific crosses. Interestingly, the early stages of pollen capture occurred irrespective of self-incompatibility or unilateral incompatibility responses in play, but did not typically proceed beyond the pollen adhesion stage (Luu et al., 1997a, 1998).
Pollen and stigma components required for pollen capture and adhesion
The early stages of pollen capture and adhesion involve the exine (Gaude and Dumas, 1984; Zinkl et al., 1999), which is the highly sculptured outer wall of the pollen grain, composed primarily of sporopollenin (reviewed in Piffanelli et al., 1998). Formation of the exine is essential for pollen grain integrity as mutants with severe exine defects also display reduced pollen viability (Paxson-Sowders et al., 2001; Guan et al., 2008). However, exine patterning is less critical as a number of mutants with disrupted exine patterning have been found to be viable (Nishikawa et al., 2005; Ariizumi et al., 2008; Suzuki et al., 2008; Dobritsa et al., 2009,a). Interestingly, the exine was found to be the only component required for the initial step of A. thaliana pollen capture onto the A. thaliana stigma (Zinkl et al., 1999), and mutant pollen grains with malformed exines have reduced adhesion to the stigmatic surface (Zinkl and Preuss, 2000; Nishikawa et al., 2005; Dobritsa et al., 2009,a). The chemical basis for this exine specificity is not yet well understood; however, insight into this specificity may emerge as the chemical nature of sporopollenin becomes better known. Through the analysis of A. thaliana mutant pollen grains that lack the exine layer, recent progress has been made in identifying the candidate biosynthetic enzymes required for exine/sporopollenin production (Morant et al., 2007; de Azevedo Souza et al., 2009; Dobritsa et al., 2009b).
Following the initial adhesive interaction, the subsequent stronger interactions between the pollen grain and stigmatic papilla require proteins and lipids from both surfaces (reviewed in Roberts et al., 1980; Dickinson et al., 2000). On the pollen side is the requirement for the pollen coat that is deposited in the interstices of the exine as shown in B. oleracea (Stead et al., 1980; Elleman and Dickinson, 1996). Lipids are the main component of the pollen coat followed by proteins, which include oleosin-like proteins, enzymes, and small pollen coat proteins (PCPs) (Doughty et al., 1993; Stephenson et al., 1997; Mayfield et al., 2001; Murphy, 2006). On the stigmatic side, early studies on Brassica spp. showed that the papillae have a waxy cuticle covered with the pellicle, a thin proteinaceous layer, which is required for pollen adhesion (Mattson et al., 1974; Stead et al., 1980; Zuberi and Dickinson, 1985,b; Elleman et al., 1992; Elleman and Dickinson, 1996). The proteins and lipids in the pollen coat and on the surface of the stigmatic papillae intermix (termed foot formation) in a process that is essential for the acceptance of compatible pollen (Elleman and Dickinson, 1990; Kandasamy et al., 1994). Poor adhesion at this stage was observed in A. thaliana mutants lacking the pollen coat and in B. oleracea when the pollen coat was removed (Preuss et al., 1993; Luu et al., 1997,a; Zinkl et al., 1999).
Cell–cell communication following pollen adhesion
One of the functions of the foot formation is likely to bring the pollen and stigmatic signalling proteins together for the putative ‘family-wide’ pollen recognition system (Fig. 2). Candidates for this recognition process are two stigma-specific proteins, the Brassica S-locus glycoprotein (SLG) and the Brassica S-locus related-1 (SLR1) proteins, which have been implicated in pollen adhesion. Stigmas from B. napus plants, with antisense suppressed SLR1, displayed reduced pollen adhesion (Luu et al., 1997,b, 1999). Furthermore, blocking either SLG or SLR1 by treating B. oleracea stigmas with their respective antibodies also reduced pollen adhesion (Luu et al., 1999). SLG was first identified as a candidate S-locus gene for Brassica self-incompatibility, but is not essential for this trait (Nasrallah et al., 1985; Takasaki et al., 2000; Silva et al., 2001). SLR1 was identified by its sequence similarity to SLG (Isogai et al., 1988; Lalonde et al., 1989; Trick and Flavell, 1989; Watanabe et al., 1992). Interestingly, SLG and SLR1 are secreted stigmatic glycoproteins with sequence similarity to the extracellular domains of the large S-domain receptor kinase family (Takayama et al., 1987; Isogai et al., 1988; Shiu and Bleecker, 2003). Thus, this raises the question of whether they are functioning alongside a related receptor kinase. In keeping with this, both SLG and SLR1 have been found to bind to the small pollen coat proteins, PCP-A1 and SLR1-BP, respectively (Doughty et al., 1998; Takayama et al., 2000), and these interactions are proposed to mediate pollen adhesion. With pollen adhesion and foot formation, a hydrophilic environment is created for pollen hydration, and this would also allow the small PCPs to freely pass to the stigmatic surface to interact with SLR1, SLG, and perhaps other stigmatic receptors to mobilize the next steps.
Similar interactions occur at this stage following a self-incompatible pollination (Fig. 2). There are two key regulators of this response, the small pollen coat protein, S-locus cysteine rich/S-locus protein 11 (SCR/SP11), and the stigma-localized S Receptor Kinase (SRK). First isolated in Brassica spp., these proteins are encoded by two tightly-linked and co-evolved polymorphic genes (reviewed in Watanabe et al., 2008). The linked SCR/SP11 and SRK alleles are referred to as S haplotypes, and pollen rejection occurs between plants sharing the same S haplotype. SCR and SRK genes have also been identified in other self-incompatible Brassicaceae spp. and characterized in more detail for A. lyrata (reviewed in Sherman-Broyles and Nasrallah, 2008). Following the attachment of ‘self’ pollen to the Brassica stigma, SCR/SP11 binds to the membrane-localized SRK, and this receptor–ligand interaction activates SRK (Kachroo et al., 2001; Takayama et al., 2001; Shimosato et al., 2007). The SRK signalling pathway is then activated in the stigmatic papilla leading to rejection of the ‘self’ pollen (reviewed in Samuel et al., 2008; Haasen and Goring, 2010). The intracellular signalling pathway includes the Brassica M Locus Protein Kinase (MLPK), a plasma membrane localized protein kinase, which interacts with and is phosphorylated by SRK. MLPK is required for the self-incompatibility response and is proposed to function in a complex with SRK to activate downstream signalling proteins (Murase et al., 2004; Kakita et al., 2007,a, b). The next step in the pathway is proposed to be the ‘activation’ of the Brassica Arm-Repeat Containing-1 (ARC1) E3 ubiquitin ligase. ARC1 binds to the activated SRK kinase domain and is required downstream of SRK for the self-incompatibility response (Gu et al., 1998; Stone et al., 1999, 2003). ARC1, as a functional E3 ligase, is proposed to inactivate factors in the stigmatic papilla that are normally required to accept compatible pollen. Recently, one factor, B. napus Exo70A1, has been identified as an ARC1 target (Samuel et al., 2009). The result of the activated self-incompatibility response is that pollen is inhibited as described further in the following sections.
The unilateral incompatibility response, described above with interspecific crosses, suggests that there are some interactions occurring in the stigma between the proteins promoting the self-incompatibility response and the ‘family-wide’ compatible pollen recognition system. With related signalling proteins involved, perhaps there are competitive binding interactions taking place between SRK, which promotes ‘self’ pollen rejection, and those receptors which positively regulate pollen acceptance. However, Kandasamy et al. (1994) reported that a Brassica line with an SRK mutation was still able to reject Arabidopsis pollen, suggesting that SRK is not required for the unilateral incompatibility response. It is important to note that surveys for unilateral incompatibility found several exceptions to the rules of unilateral incompatibility suggesting that this trait may be quite complex (Sampson, 1962; Hiscock and Dickinson, 1993; Luu et al., 1998). A more complete picture of the molecular interactions occurring as part of the unilateral incompatibility response will clearly require a broader understanding of the initial signalling in the stigma for the ‘family-wide’ compatible pollen recognition, at both the protein and genetic levels.
Pollen hydration, following adhesion to the stigmatic papillae
The pollen hydration phase is an important step in the compatible pollen response as it allows the quiescent, desiccated pollen grain to regain its metabolic activity prior to pollen tube emergence (Fig. 2). Compared to wet stigmas on which pollen grains hydrate by default, the process of pollen grain hydration on dry stigmas is highly regulated (Zuberi and Dickinson, 1985,a; Sarker et al., 1988). Pollen hydration is one of the earliest steps blocked in a self-incompatible pollination (Dickinson and Elleman, 1985; Dickinson, 1995). The lipid and proteinaceous components of the pollen coat are essential to pollen hydration, and during pollen foot formation, there are changes to the pollen coat, termed coat conversion, that were initially observed in B. oleracea (Elleman and Dickinson, 1986). In this process, the lipids are thought to reorganize, perhaps through the actions of the lipid-binding oleosin-like proteins, to create a capillary system through which water can flow from the stigma to the pollen grain (Murphy, 2006).
The role of lipids in pollen hydration
Clues to the importance of pollen coat lipids came from A. thaliana mutants with defects in long-chain lipid metabolism such as the male sterile eceriferum (cer) mutants (Preuss et al., 1993; Hulskamp et al., 1995,a). In a strong cer6 mutant, the absence of long-chain lipids resulted in an absence of pollen coat on the pollen grain surface and the pollen failed to hydrate on the stigma. Interestingly, this defect could be rescued by high environmental humidity or the application of lipids to the stigma, both of which allowed the pollen grain to hydrate and germinate leading to successful fertilization (Preuss et al., 1993; Wolters-Arts et al., 1998). A weaker cer6 mutant and the cer1 mutant showed a reduction in the lipid droplets of the pollen coat and also failed to hydrate on the stigma, but this defect could be rescued by co-pollination with wild-type pollen (Preuss et al., 1993; Hulskamp et al., 1995,a). Further characterization of these cer mutants and their respective genes showed that CER1 was an enzyme needed for the conversion of long chain aldehydes to alkanes during the process of wax biosynthesis (Aarts et al., 1995), that CER6 is required for the production of long chain fatty acids (Fiebig et al., 2000).
In contrast to the aforementioned lipid mutants is the A. thaliana fiddlehead (fdh) mutant that showed altered cuticle composition over the entire shoot and allowed for pollen germination and pollen tube growth on the shoot epidermis, but not on cotyledons or roots (Lolle and Cheung, 1993). Intriguingly, Lolle and Cheung (1993) tested the species specificity of this and found that pollen from other Brassicaceae spp. had a similar response as A. thaliana pollen, while pollen grains from plants outside of this family adhered poorly to the fdh mutant shoot epidermis. Thus, the ‘family-wide’ recognition system seems to be intact in the shoot epidermis of the fdh mutant. When pollen grains from the cer mutants were tested for hydration on the fdh mutant shoot epidermis, no hydration was detected, indicating that the pollen coat was required for this response (Lolle et al., 1997). Lipid profiles from the fdh shoots showed an increase in long chain fatty acids relative to the wild type and, consistent with this, the FDH gene is predicted to encode a long-chain fatty acid elongase (Lolle et al., 1997; Pruitt et al., 2000). Interestingly, the application of lipids on wild-type leaves also promoted pollen germination and pollen tube growth (Wolters-Arts et al., 1998).
The pollen oleosin-like proteins have been implicated in pollen hydration through the analysis of the A. thaliana glycine-rich protein (grp)17 mutant (Mayfield and Preuss, 2000). The grp17 mutant pollen had a pollen coat that was normal in appearance, but was missing the GRP17 oleosin-domain protein. The effect of this was a significant delay in the initiation of hydration, in comparison to wild-type pollen, although once initiated, a normal rate of hydration was observed. A pollen coat enzyme has also been implicated in pollen hydration through the analysis of a T-DNA insertion mutant for the A. thaliana extracellular lipase 4 (EXL4) gene (Updegraff et al., 2009). The exl4 mutant pollen is morphologically the same as wild-type pollen grains and initiated hydration at roughly the same time as the wild-type, but at a slower rate, resulting in a significantly longer time for hydration. The exl4 mutant pollen was found to have reduced esterase activity, relative to the wild type, suggesting that EXL4 is an esterase with a role in lipid modification (Updegraff et al., 2009).
Stigmatic responses regulating pollen hydration
Recently, Exo70A1 was identified as a protein required in the stigma for pollen hydration in B. napus and A. thaliana. Either the absence or a reduction of Exo70A1 in the stigma resulted in poor pollen hydration for both A. thaliana and B. napus. In A. thaliana, an RFP-tagged B. napus Exo70A1 construct was able to rescue this defect, and, interestingly, RFP:Exo70A1 was localized to the plasma membrane in the stigmatic papillae of mature flowers (Samuel et al., 2009). Initially, Exo70A1 was isolated as a substrate for the Brassica ARC1 E3 ubiquitin ligase, and the reduced pollen hydration associated with the loss of Exo70A1 fits with ARC1's role in the inhibition of compatibility factors, such as Exo70A1, to promote the self-incompatibility response. Consistent with this, the overexpression of RFP:Exo70A1 in B. napus stigmas was able to partially overcome the self-incompatibility response, favouring self-compatibility (Samuel et al., 2009). Exo70A1 is a putative subunit of the plant exocyst complex proposed to be involved in the tethering of post-Golgi secretory vesicles to the plasma membrane (reviewed in He and Guo, 2009; Zarsky et al., 2009). Thus, Exo70A1, in its role with the exocyst complex, may be involved in tethering secretory vesicles to the stigmatic papilla plasma membrane at the pollen contact site to deliver other stigmatic factors required for pollen hydration. Vesicle-like structures have been observed in B. oleracea stigmatic papillae following treatment with pollen coat (Elleman and Dickinson, 1996).
The signalling events that regulate the stigmatic responses to compatible pollen at this stage involve calcium. Following pollination, real-time imaging of calcium levels in the A. thaliana stigmatic papillae led to the discovery of three cytosolic calcium increases in the apical region of the papilla: the first following pollen hydration, the second increase prior to pollen germination, and the third increase when pollen tube penetration of the stigmatic cell wall occurred (Iwano et al., 2004). In B. rapa, changes in the actin cytoskeleton were also documented following pollination (Iwano et al., 2007). The application of either compatible pollen grains or pollen coat, induced actin polymerization in the apical region of the stigmatic papilla. This resulted in the increased formation of actin bundles at the apical tip at a time when the pollen grains were undergoing hydration (Iwano et al., 2007), an observation consistent with the idea of secretory vesicles being delivered along the actin cytoskeleton to the apical tip/pollen contact site for exocyst tethering and membrane fusion (He and Guo, 2009). In addition, changes in the tubular vacuolar network in the apical region of the stigmatic papilla were observed with the large central vacuole orienting towards the pollen contact site with compatible pollen, perhaps to promote pollen hydration (Iwano et al., 2007). By contrast, the application of self-incompatible pollen grains or pollen coat was associated with a decrease in actin filaments in the apical region and a more disorganized appearance of the tubular vacuolar network in the stigmatic papillae (Iwano et al., 2007).
Pollen germination and pollen tube growth through the stigmatic surface
Once the pollen grain has successfully hydrated, the pollen germinates and a pollen tube emerges (Fig. 2). The pollen tube typically grows through the foot to penetrate the stigmatic cuticle and then enters the outer layer of the stigmatic cell wall (Elleman et al., 1992; Kandasamy et al., 1994). For B. oleracea, the pollen tube grows through the inner and outer layers of the cell wall (Elleman et al., 1992). A. thaliana pollen tubes also grow through the two layers of the stigmatic cell wall, but were observed to grow between the cell wall and the plasma membrane as well (Elleman et al., 1992; Kandasamy et al., 1994). The invasion of the pollen tube through the stigmatic cuticle and cell wall would be predicted to require enzyme modification of these layers, to allow further pollen tube growth. Searches for cutinases or esterases that could break down the stigmatic cuticle have uncovered esterase activities in both Brassica pollen and the stigma extracts, and treatment of stigmas with a serine esterase inhibitor blocked pollen tube penetration (Hiscock et al., 1994, 2002; Lavithis and Bhalla, 1995). The stigmatic cell wall at the pollen contact point has been observed to be expanded, presumably due to the action of cell wall-modifying enzymes (Elleman and Dickinson, 1990; Kandasamy et al., 1994). Similarly, cell wall-modifying enzymes such as polygalacturonases and pectin esterases have also been identified in Brassica pollen (Kim et al., 1996; Dearnaley and Daggard, 2001). The initial expansion of the stigmatic cell wall, however, is more likely to be due to enzymes secreted by the stigmatic papilla and is consistent with observation by Elleman and Dickinson (1996) that ER, Golgi, and vesicle-like structures are associated with the sites of stigmatic cell wall expansion.
Exo70A1 is also required in the stigma for the penetration of compatible pollen tubes. When B. napus stigmas with reduced levels of Exo70A1 mRNA were pollinated with compatible pollen, very little seed set was observed and most pollen tubes failed to penetrate the stigmatic barrier, although pollen tubes could be seen growing over the surface of papillae cells unable to penetrate the cuticle or cell wall (Samuel et al., 2009). Similar to the role of the exocyst in pollen hydration (described in the previous section), the tethering of secretory vesicles by the exocyst complex to the stigmatic papilla plasma membrane at the pollen contact site may be required for the delivery of enzymes required for pollen tube penetration through the stigmatic cuticle and cell wall. Once this process is initiated, the pollen tube probably produces its own cell-wall modifying enzymes and is able to partially support its continued growth through the cell wall layers of the stigmatic papillae.
Pollen tube guidance to the female gametophyte
When pollen tubes emerge at the base of the stigmatic papillae, they grow intercellularly down to the style and then through the transmitting tissue of the style and septum (a central tissue that runs to the base of the ovary) (Hill and Lord, 1987; Lennon et al., 1998). Once the pollen tube emerges from the septum, it grows over the surface of the septum to a funiculus (which attaches the ovule to the septum). The pollen tube then grows along the funicular surface to the micropylar opening of the ovule where it enters to release the sperm cells (Fig. 1) (reviewed in Yadegari and Drews, 2004). The transmitting tissue is a specialized column of cells producing extracellular matrix (ECM) material through which the pollen tubes navigate en route to the ovules. It provides chemical gradients and nutrients for pollen tube guidance and growth, and proper transmitting tract formation is essential for efficient pollen tube guidance (reviewed in Crawford and Yanofsky, 2008). For example, the No Transmitting Tract (NTT) gene in A. thaliana is required for normal transmitting tract development, and in ntt mutants, pollen tubes are no longer restricted to growth through the septum. In addition, pollen tube elongation was slower and more jagged in this mutant, in comparison to the wild type (Crawford et al., 2007). Despite the abnormal transmitting tract in the ntt mutants, pollen tubes were able to grow through the style and fertilize the ovules at the apex of the ovary. This shows that some long-range pollen tube guidance systems are still functional, since the pollen tube can be directed to the funiculus from what remains of the transmitting tract (Crawford et al., 2007).
Pistil factors regulating pollen tube guidance
In recent years, it has become more clear that chemo-attractant gradients in the pistil play an important role in guiding pollen tubes to the ovules (Fig. 3) (reviewed in Johnson and Lord, 2006; Crawford and Yanofsky, 2008; Higashiyama and Hamamura, 2008). One of the first molecules proposed to guide pollen tubes was γ-amino butyric acid (GABA) that was identified through the analysis of the A. thaliana pollen-pistil (pop)2 mutant (Wilhelmi and Preuss, 1996; Palanivelu et al., 2003). The pop2 mutant displayed abnormal pollen tube guidance. The pop2 pollen tubes were less able to find the micropyle and frequently displayed an atypical behaviour with more than one pollen tube present on a single funiculus. This defective pollen tube guidance required both the pollen and pistil to carry the recessive pop2 mutation, as wild-type pollen tubes behaved normally on pop2 pistils (Wilhelmi and Preuss, 1996). The pop2 mutant was found to lack a functional GABA transaminase, which led to excess levels of GABA in the pistil. In wild-type pistils, GABA was found to be present as a gradient, starting from the stigma and increasing in concentration to the inner integument of the ovule, and this gradient was proposed to guide the growing pollen tube (Palanivelu et al., 2003). Further advances in the understanding of chemo-attractants were provided by the discovery of chemocyanin in the lily system (Kim et al., 2003), followed by the identification of the structurally-related plantacyanin in A. thaliana (Dong et al., 2005). Both peptides exist in increasing gradients along the style, and transmission tract leading to the ovary. When plantacyanin was overproduced in A. thaliana pistils, wild-type pollen tubes had difficulties at the earliest stages of pollen tube guidance, circling the stigmatic papillae and even growing away from the style (Dong et al., 2005).
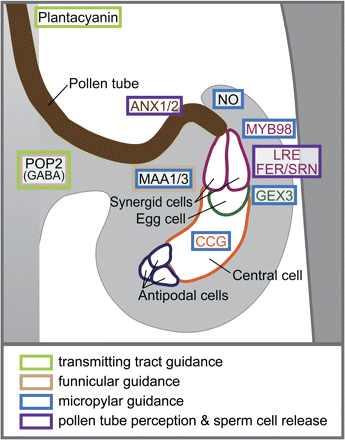
Model of pollen tube guidance to the female gametophyte in A. thaliana. An illustration of a pollen tube growing to an ovule is shown, with the guidance cues and genes that are proposed to regulate pollen tube guidance and perception overlaid on this diagram. If expression patterns are known, gene names are coloured to match the cells where they are expressed. Coloured boxes indicate steps that are disrupted in mutants. Please see the text for further details and references.
During the course of the last decade, the understanding of the role of the female gametophyte in producing pollen tube guidance cues has increased immensely with the discovery of funicular and micropylar guidance systems (Fig. 3). Initially, genetic screens for pollen guidance defects uncovered mutants in which the absence of the female gametophyte results in impaired guidance of pollen tubes to the ovule (Hulskamp et al., 1995,a; Ray et al., 1997). The novel Gamete-Expressed (GEX)3 gene, which is expressed in the egg cell, is one factor that acts at this stage. Reduced GEX3 expression in the pistil resulted in wild-type pollen tubes being unable to locate the micropyle following growth along the funiculus (Alandete-Saez et al., 2008). A comparable phenotype was also observed in the A. thaliana magatama1 (maa1) and magatama3 (maa3) mutants, in which female gametophyte development was delayed, and pollen tubes were unable to find the micropyle (Shimizu and Okada, 2000). Interestingly, Shimizu and Okada (2000) found similar phenotypes when examining the path of pollen tubes following interspecific crosses with wild-type pistils. When either B. napus or Orychophragmus violaceus pollen was applied to A. thaliana pistils, the pollen tubes grew to the septum, but very few were guided to the ovules, with some pollen tubes mimicking the maa phenotype. Interspecific pollen guidance to the micropyle was also observed to be inefficient in a study with Torenia fournieri (Higashiyama et al., 2006). Thus, interspecific barriers may exist at the level of the micropylar guidance system to prevent unproductive fertilization events. Another phenotype observed at high frequency in the A. thaliana maa mutants was two pollen tubes approaching a single ovule. This suggested that repulsive signals from the ovule to deter the approach of more than one pollen tube at the funiculus were disrupted (Shimizu and Okada, 2000). Recently, the MAA3 gene was cloned and predicted to encode a helicase with a role in RNA processing and metabolism (Shimizu et al., 2008).
In searching for repulsive signals that would restrict more than one pollen tube from growing on the funiculus, nitric oxide has been suggested as a candidate for this signal (Crawford and Yanofsky, 2008) on account of its ability to cause a sharp turn in growing lily pollen tubes (Prado et al., 2004). More recently, support for the role of nitric oxide in micropylar guidance has emerged in a study with the A. thaliana nitric oxide synthase (nos)1 mutant that is defective in nitric oxide production and displays reduced fertility (Guo et al., 2003; Prado et al., 2008). Wild-type pollen tubes were able to grow in proximity to nos1 ovules, but became deformed as they approached the micropyle (Prado et al., 2008). In addition, staining for nitric oxide in wild-type ovules showed an asymmetric distribution around the micropyle, perhaps indicating a function in funnelling pollen tube at the micropyle (Prado et al., 2008).
The analysis of mutants defective in micropylar guidance established the presence of this guidance system, but the question then arose as to which female gametophyte cells were the source of this signal. At maturity, the female gametophyte is composed of two synergid cells, the egg cell, a central cell and three antipodal cells (reviewed in Punwani and Drews, 2008). Laser ablation studies in Torenia fournieri, in which different female gametophytic cells were systematically destroyed, provided compelling evidence for a micropylar guidance signal originating from the synergid cells (Higashiyama et al., 2001). More recently, the attractant signal being secreted from the T. fournieri synergid cells was identified as a group of proteins, termed the LURE proteins, which belong to the defensin-like subgroup of cysteine-rich polypeptides (Okuda et al., 2009). Interestingly, the Brassica SCR/SP11 protein, which functions as a pollen ligand in the self-incompatibility response, also belongs to this defensin-like family of proteins (Mishima et al., 2003).
In A. thaliana, support for the role of synergid cells in guiding pollen tubes to the micropyle came from the analysis of the myb98 mutant (Kasahara et al., 2005). This mutant displayed normal female gametophyte development, with the exception of the synergid cells. As a result, pollen tubes would grow to the funiculus, but were unable to find the micropyle, suggesting that the affected synergid cells were not producing the required signal for this guidance step (Kasahara et al., 2005). MYB98 is a transcriptional regulator expressed specifically in the synergid cells where it functions to activate the expression of genes required for both pollen tube guidance and the formation of the synergid cell filiform apparatus (Punwani et al., 2007). The MYB98-regulated genes included predicted genes for small cysteine-rich proteins, similar to the T. fournieri LURE proteins (Jones-Rhoades et al., 2007; Punwani et al., 2007, 2008). Another transcriptional regulator, Central Cell Guidance (CCG), expressed in the central cell of the ovule, has also been found to be necessary for pollen tube guidance to the micropyle in A. thaliana (Chen et al., 2007).
Pollen tube perception in the ovule
Once the pollen tube enters the micropyle, the final stage is pollen tube perception, where the pollen tube penetrates a synergid cell and bursts to release the two sperm cells for fertilization (Fig. 3). In the A. thaliana feronia (fer)/sirène (srn) mutant, the pollen tubes enter the micropyle but fail to stop growing and are unable to rupture which results in the formation of pollen tube coils within female gametophyte (Huck et al., 2003; Rotman et al., 2003). The FER/SRN gene encodes a receptor kinase belonging to the CrRLK1 subfamily of receptor kinases and is expressed in the synergid cell in A. thaliana (Escobar-Restrepo et al., 2007; Hematy and Hofte, 2008). Interestingly, Escobar-Restrepo et al. (2007) also tested interspecific crosses between A. thaliana and two other Brassicaceae species, A. lyrata and Cardamine flexuosa, and observed interspecific barriers at the level of pollen tube perception. When the more closely related A. lyrata pollen was applied, roughly half of the pollen tubes successfully fertilized A. thaliana ovules, while most of the remaining A. lyrata pollen tubes displayed the overgrown fer/srn-like phenotype. For the more distantly related C. flexuosa pollen, many fewer A. thaliana ovules attracted C. flexuosa pollen tubes, and the C. flexuosa pollen tubes either coiled outside the micropyles, stopped just after entering the micropyles, or entered the micropyles and continued to grow in the fer/srn-like phenotype in the wild-type A. thaliana ovules (Escobar-Restrepo et al., 2007). The A. thaliana LORELEI (LRE) gene, which encodes a putative glucosylphosphatidylinositol-anchored protein, is also expressed in the synergid cells, and the lre female gametophyte mutant displays impaired sperm cell release, similar to the fer/srn mutant (Capron et al., 2008).
Pollen factors required for pollen tube guidance and perception
While the discussion has largely focused on pistil factors required for compatible pollinations, pollen tube guidance, and fertilization, there is also a basic requirement of intact cellular processes within the pollen tube to driving rapid polar growth (reviewed in Cheung and Wu, 2008; Moscatelli and Idilli, 2009). In addition, the pollen tubes need to perceive the guidance cues within the pistil to direct growth towards the female gametophyte. Therefore, genetic screens have also focused on identifying male mutants which display disrupted pollen tube guidance to the ovule as an approach to finding these predicted pollen tube receptors (Johnson et al., 2004; Boavida et al., 2009). For example, the Lycopersicon esculentum pollen-specific receptor kinase, LePRK2, is required for pollen germination and tube growth, and plays a role in responding to growth-promoting signals from the pistil (Zhang et al., 2008). A candidate for this growth-promoting signal is LeSTIG1, a small cysteine-rich pistil protein, which binds to the extracellular domain of LePRK2 and promotes pollen tube growth in vitro (Tang et al., 2004). In A. thaliana, the characterization of the FER/SRN receptor kinase in the synergid cells has recently led to the identification of two closely related members functioning in the pollen tube to regulate sperm cell release (Boisson-Dernier et al., 2009; Miyazaki et al., 2009). The ANXUR1 (ANX1) and ANXUR2 (ANX2) genes are expressed at highest levels in the pollen, and double anx1/anx2 mutants produce pollen tubes which rupture prematurely. Thus, ANX1 and ANX2 are proposed to function in the pollen tube to co-ordinate the timing of pollen tube rupture and release of the sperm cells, in conjunction with the FER/SRN receptor kinase signalling in the synergid cells (Boisson-Dernier et al., 2009; Miyazaki et al., 2009). Given the late stage at which these receptor kinases function, there are presumably additional pollen tube receptors required for sensing guidance cues in the transmitting tissue. The genetic screens for pollen tube guidance defects have identified male mutants with disrupted pollen tube guidance at earlier stages, and look promising for uncovering new players in this process (Johnson et al., 2004; Boavida et al., 2009).
Conclusions
At the stigmatic surface of Brassicaceae pistils, there are complex pollen recognition signalling systems at play to determine whether a pollen grain should be accepted. There is evidence to support a ‘family-wide’ pollen recognition system that allows for interspecific and intergeneric crosses; however, some preference for species-specific pollen may occur by different initial binding affinities of pollen grains (Zinkl et al., 1999; Hiscock and Dickinson, 1993). As such, the identification of all protein and lipid factors contributing to pollen adhesion and hydration efficiency will be essential to understanding these interactions more fully. In addition, the self-incompatibility system is intrinsically linked to the compatible pollen–pistil interactions and functions to override the compatible pollen responses when activated (Samuel et al., 2009). Thus, increasing our understanding of compatible pollen–pistil interactions will provide an insight into how the self-incompatibility pathway functions in the stigmatic papillae to block what would otherwise be recognized as compatible pollen.
Once the pollen tube has penetrated the stigmatic surface and continues to grow towards the ovule, there appear to be other recognition systems in play for species selectivity. Little support currently exists for a species-specific filter in the septum; however, species specificity has been proposed in the form of variable concentrations of chemo-attractant molecules and the differential abilities of pollen tubes from different species to detect such compounds (Johnson and Lord, 2006). A family-wide comparison of chemo-attractant molecules would be needed to evaluate such an hypothesis. Nevertheless, there is evidence for other layers of species discrimination in regulating the final stages of pollen tube guidance. This includes the micropylar guidance system that functions most efficiently with species-specific pollen tubes (Shimizu and Okada, 2000). In addition, barriers in the female gametophyte act at the pollen tube perception stage to prevent fertilization between more distantly related species in the Brassicaceae (Escobar-Restrepo et al., 2007). Finally, while not discussed in this review, there are reproductive barriers taking place post-fertilization, such as with interploidy crosses, which lead to non-viable embryos (for a review see Dumas and Rogowsky, 2008).
We thank members of the Goring laboratory for critical reading of the manuscript. LAC is supported by a graduate scholarship from the Natural Sciences and Engineering Research Council of Canada (NSERC), and research in the laboratory of DRG is supported by grants from NSERC and a Canada Research Chair to DRG.

{kind=link}
{kind=link}
{kind=link}
Comments